USMP - Filial Norte

Facultad de Medicina Humana
miércoles, 5 de noviembre de 2008
¿Nuestra Conducta está Escrita en Nuestros Genes?
El presente es un resumen a modo de comentario acerca del siguiente artículo científico: Enlace al artículo Original
A la pregunta ¿nuestra conducta está escrita en nuestros genes?, la respuesta se deja pensar por un momento…
El artículo hace referencia a un nuevo estudio que demostró el papel que desempeña un solo gen en el comportamiento sexual específico de la mosca de la fruta Drosophila melanogaster. Este estudio se enfocó en un gen llamado “fruitless”. El producto del ARN mensajero de este gen (ver FIGURA 4) codifica un factor de transcripción que determina dos formas específicas para el sexo, una de ellas es única para las moscas macho y la otra lo es para las moscas hembra.
Demir y Dickson (los investigadores) emplearon manipulación genética para producir, anatómicamente, moscas hembras que incluyeran sólo la forma masculina del gen (ver FIGURA 4). Las moscas hembra resultantes exhibieron cortejo y conducta de apareamiento hacia las hembras. Mientras que los estudios previos han mostrado que la forma masculina del gen fruitless es necesaria para el cortejo masculino, el nuevo estudio señala que este gen es suficiente para producir esa conducta, aun en las hembras.
Luego, se discuten las preguntas: ¿Qué otras formas de conducta con tales manifestaciones complejas podrían probar tener un origen tan simple? ¿Es concebible que las conductas complejas en humanos pueden estar determinadas por un solo gen?... Las dejo sin comentar para provocarle curiosidad y le invito a que continúe la lectura en el artículo original:
Enlace al artículo Original
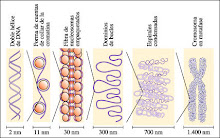
El Cromosoma Eucariótico
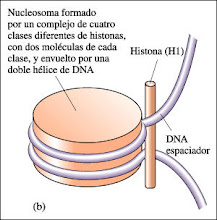
El DNA eucariótico siempre está asociado con proteínas, que constituyen más de la mitad del peso del cromosoma. La mayoría de estas proteínas son histonas, moléculas relativamente pequeñas con carga positiva. La molécula de DNA se envuelve alrededor de núcleos formados por ocho moléculas de histonas, para formar nucleosomas, las unidades de empaquetamiento básico del DNA de los eucariotas. Los nucleosomas se empaquetan unos sobre otros formando una estructura más condensada -la fibra de 30 nanómetros- que se encuentra tanto en la cromatina en la etapa de interfase como en los cromosomas que entran en mitosis. El DNA puede tomar in vitro la forma de B-DNA (la hélice dextrógira descrita por Watson y Crick), A-DNA (una hélice dextrógira menos fuertemente enrollada) o Z-DNA (una hélice levógira). Las distintas formas del DNA podrían coexistir in vivo, con distintas funciones biológicas.
La distancia entre los nucleosomas es entre 10 y 11 nanómetros y el diámetro de cada cuenta es aproximadamente de 7 nanómetros. El centro de cada nucleosoma está compuesto por alrededor de 140 pares de bases de DNA y un conjunto de ocho moléculas de histona. La cadena que une los núcleos de los nucleosomas contiene otros 30 ó 60 pares de bases.
La replicación del DNA en los eucariotas es igual, en principio, a la replicación del DNA de los procariotas. Los nucleótidos en la forma de trifosfatos se acoplan a lo largo de una cadena molde de DNA en la forma semiconservativa. Como en los procariotas, en la síntesis de las cadenas complementarias, las DNA polimerasas operan solamente en una dirección''; la cadena complementaria a la otra hebra madre ''se sintetiza como una serie de fragmentos de Okazaki, que luego se unen por la acción de la enzima DNA ligasa.
En el cromosoma procariótico circular, comparativamente pequeño, la replicación comienza en un único origen y procede bidireccionalmente a lo largo de dos horquillas de replicación.
Las histonas ensambladas en los nucleosomas raramente se separan del DNA, por lo tanto, a medida que la horquilla de replicación avanza debe, de alguna manera, pasar a través de los nucleosomas parentales. Se cree que cada nucleosoma se desenrolla transitoriamente, permitiendo que la DNA polimerasa copie el DNA nucleosomal desenrollado. El DNA recién sintetizado hereda parte de las viejas histonas pero requiere también de nuevas histonas para empaquetarse y formar la cromatina. En cuanto se sintetiza, el DNA eucariótico forma los nucleosomas y se asocia también con otras proteínas compactándose en los niveles superiores de condensación.
Material original en: curtisbiologia.com
Extraído de: Biología de Curtis & Barnes
Introducción

Con la ayuda creciente de las herramientas de la tecnología del DNA recombinante, progresaron los estudios de genética molecular de los eucariotas y la distancia entre los eucariotas y los procariotas, en vez de disminuir, se fue ampliando.
Un organismo multicelular usualmente inicia su vida en forma de huevo fecundado, el cigoto. El cigoto se divide repetidamente produciendo muchas células que se diferencian y cada tipo celular comienza a producir proteínas característicamente diferentes que lo distinguen de otros tipos de células. A su vez, un mismo tipo celular puede producir variantes de las proteínas que sintetiza en distintas etapas del desarrollo del organismo. Sin embargo, toda la información genética originalmente presente en el cigoto también está presente en cada célula diploide del organismo. Resulta claro que la diferenciación de las células de un organismo multicelular depende de la inactivación de ciertos grupos de genes y de la activación de otros, es decir, de una regulación de la expresión.
La expresión de los genes puede ser regulada en distintas etapas del camino que conduce desde el DNA a las proteínas. Las investigaciones en eucariotas han demostrado que, como en el caso de los procariotas, el genoma eucariótico contiene un sorprendente arreglo de elementos genéticos móviles. El análisis del cromosoma eucariótico ha revelado que éste también está sujeto a reordenamientos, deleciones y adiciones.
En ocasiones, las células escapan a los factores que regulan el crecimiento celular normal. En consecuencia, las células se multiplican sin control, amontonándose, invadiendo y destruyendo otros tejidos. En estos casos se produce el cáncer. Hay varias evidencias que han relacionado el desarrollo del cáncer con cambios en el material genético. Los virus pueden provocar cambios en la constitución genética de la célula y algunos pueden causar cáncer. Más aun, todos los virus conocidos causantes de cáncer son virus que introducen información en los cromosomas de las células hospedadoras. Estos incluyen tanto virus con genoma de DNA como retrovirus con genoma de RNA. El descubrimiento del papel de la transcriptasa inversa forjó el eslabón crucial entre los retrovirus y los cromosomas de las células eucarióticas.
El mayor anhelo relativo a las aplicaciones basadas en la tecnología del DNA recombinante es que en algún tiempo futuro sea posible corregir defectos genéticos sustituyendo genes "malos " por genes "buenos". Esta es una tarea enormemente compleja. Requiere primero la preparación de un gen que será captado por una célula eucariótica, incorporado a un cromosoma, y luego expresado, pero esto es sólo el comienzo. El nuevo gen debe establecerse en un gran número de células del tipo apropiado y quedar sujeto a los complicados, y todavía grandemente desconocidos, controles del gen normal.
Se han desarrollado técnicas que permiten la transferencia de genes a células eucarióticas. Las células se hacen crecer en cultivo en tubos de ensayo y se transfieren a óvulos fecundados de ratón y de otras especies de mamíferos y a embriones de Drosophila.
En cada uno de estos casos, genes extraños han sido incorporados y expresados en el nuevo hospedador. En los últimos años también se desarrollaron técnicas que permiten obtener copias idénticas de organismos, los clones.
Material original en: curtisbiologia.com
Extraído de: Biología de Curtis & Barnes
Suscribirse a:
Entradas (Atom)